

Supplier:
BioLegendCat no: 585802
Recombinant Human TGF-beta3 (carrier-free)
Prices direct from BioLegend
Quick response times
Exclusive Absave savings/discounts
SPECIFICATIONS
Catalog Number
585802
Size
5 microg
Gene Id
7043
Storage Temp
-20 C or -70 C
Shipping Temp
Blue Ice
Storage Buffer
Unopened vial can be stored at 4C for three months, at -20C for six months, or at -70C for one year. For maximum results, quick spin vial prior to opening. Stock solutions should be prepared at no less than 10 microg/mL in sterile buffer (PBS, HPBS, DPBS, and EBSS) containing carrier protein such as 1% BSA or HAS or 10 % FBS. After dilution, the cytokine can be stored at 4C for one month or from -20C to -70C for up to 3 months. Avoid repeated freeze/thaw cycles.
Alternative Names
Transforming growth factor beta 3 (TGF b3), TGFB3, TGF beta 3,ARVD
Regulatory Status
RUO
SUPPLIER INFO

Email:
info@biolegend.com





More from BioLegend

Applications
FC, ICC
Hosts
Mouse
Reactivities
Hum
Applications
FC, ICC
Hosts
Mouse
Reactivities
Hum
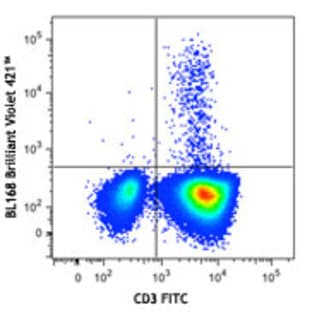
Applications
FC, ICC
Hosts
Mouse
Reactivities
Hum

Applications
FC, ICC
Hosts
Mouse
Reactivities
Hum
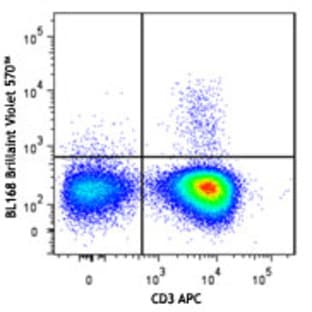
Applications
FC, ICC
Hosts
Mouse
Reactivities
Hum
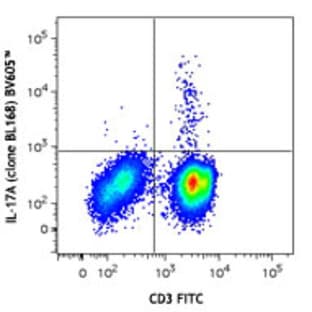
Applications
FC, ICC
Hosts
Mouse
Reactivities
Hum
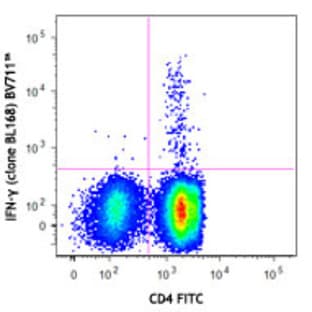
Applications
FC, ICC
Hosts
Mouse
Reactivities
Hum
Latest promotions

Buy any polyclonal or monoclonal antibody from our extensive range of pre-made antibodies and for a limited time only receive a $50 discount!(T&C apply:...

New brilliant antibodies, and new lower prices!For flow cytometry reagents in general, \"bright is better.\" The violet-excitable BD Horizon™ BV421 and...

Did your supplier increase the price of Fetal Bovine Serum? Did they substitute the US Origin with USDA? Well say no more! Innovative Research is still...

We're so sure that you'll prefer Cayman Assay kits over your present brand that we're willing to give you a free assay kit to prove it!

For the past decade scientists have extensively used ATS secondary toxin conjugates to make their own targeted toxins for in vitro use.The ability to combine...

10% Discount on 2 Rabbit Polyclonal Antibody Service. With over 20 years experience, SDIX has developed into the premier US custom antibody producer,...

Bulk Cytokines with Custom Vialing.20 - 50% off cytokines, growth factors, chemokines and more...For a limited time Cell Sciences is offering substantial...
Are you planning to have a customised antibody made for your research?Since 2000, Everest has been producing a catalog containing thousands of affinity...
Top suppliers

Agrisera AB
11 products

Biotrend

Biosensis
969 products

ABBIOTEC
3011 products

SDIX
1 products

Spring Bioscience
2291 products

Cell Signaling Technology
4976 products

Rockland Immunochemicals, Inc.
7592 products

Boster Immunoleader
1533 products

OriGene Technologies Inc.
5281 products
Maine Biotechnology Services
227 products

BD (Becton, Dickinson and Company)
1 products
ABNOVA CORPORATION
Randox Life Sciences
1502 products
